RESEARCH AND DEVELOPMENT
RESEARCH AND DEVELOPMENT

Neuroscience
Advancing the next generation of treatments for diseases of the central nervous system (CNS)
Arvinas is pioneering the discovery and development of novel PROTAC (PROteolysis-TArgeted Chimera) protein degraders as potential disease modifying therapies for neurologic disease with high unmet need. Many CNS targets have been considered “undruggable,” and are further complicated by the difficulty of delivering therapeutic agents (such as genomic modalities and antibodies) into the brain. Arvinas is exploring a new and exciting territory, developing blood-brain barrier penetrant PROTACs against target proteins that are genetically implicated in neurodegenerative diseases, such as progressive supranuclear palsy (PSP), Parkinson’s disease, Huntington’s disease, and Alzheimer’s disease.

Propelling Research for Treating Diseases of the Central Nervous System
Many CNS targets have long been considered “undruggable” due to both scaffolding functions and the lack of catalytic binding sites required for typical small molecule inhibitors, and the difficulty of delivering genomic therapeutic agents into the brain. As a result, there remains a high unmet need in many neurologic diseases, including neurodegenerative disorders. Driven by our commitment to improve the lives of patients with serious diseases, Arvinas is exploring an exciting new territory with the discovery of novel brain penetrant PROTACs.
Tackling the Problem Using Differentiated Pharmacology
As we investigate how PROTACs might become future therapeutics, we believe the differentiated pharmacology has the potential to translate into improved clinical outcomes for patients in therapeutic areas with limited or ineffective treatment options.

Potential pharmacological differentiation of the investigational Arvinas PROTAC Discovery Engine:
Potential to cross the blood-brain barrier, which is a substantial challenge for monoclonal antibodies and genomic therapies.
Potential to enter cells of the CNS, degrading pathogenic proteins (such as mutant or oligomeric tau) from the inside of the cell, thus hindering both intracellular dysfunction and “prion-like” spread of pathologic protein into the extracellular space and healthy neurons.
Potential to selectively degrade mutant forms of pathologic proteins and spare healthy copy of the protein (such as for mutant huntingtin protein (mHTT)).
Degradation enables a PROTAC to disrupt the scaffolding function of a disease-causing target protein.
Iterative (often described as “catalytic”) activity, by which a single PROTAC molecule can induce the degradation of hundreds of copies of the disease-causing protein.
Directed degradation to appropriate tissues while avoiding others by cell- or tissue-restricted E3 ligases.
Oral dosing provides an additional route of administration for the targeted elimination of disease-causing proteins from the CNS.
Clinical Studies
Arvinas’ first clinical-stage PROTAC is ARV-102. ARV-102 is a novel oral PROTAC designed to cross the blood-brain barrier and target leucine-rich repeat kinase 2 (LRRK2), a large, multi-domain scaffolding kinase. Increased activity and expressions of LRRK2 causes and is linked to the pathogenesis of neurological diseases including progressive supranuclear palsy (PSP) and Parkinson’s disease.
ARV-102 is currently under investigation in Phase 1 clinical studies. The safety and effectiveness of ARV-102 have not yet been established.
About LRRK2
What is LRRK2?
LRRK2 (Leucine-rich repeat kinase 2) is a large, multi-domain scaffolding protein. Mutations and genetic variants in LRRK2 cause or are associated with an increased risk of certain neurodegenerative diseases, particularly progressive supranuclear palsy (PSP) and Parkinson’s disease (PD).
How does LRRK2 relate to neurodegenerative diseases?
Certain mutations or genetic variants can increase the activity or expression of LRRK2, leading to dysfunction in cellular processes that are critical for maintaining the health and function of both neurons, particularly dopaminergic neurons, which are involved in motor control and microglia, which control normal neuroimmune function with aging.
Why is LRRK2 an important target?
LRRK2 is an important PROTAC degradation target because of the scientific data demonstrating that scaffolding function promotes neuronal death, reducing LRRK2 rescues neuronal loss in PD models, and increased activation and expression level may contribute to PD and PSP.
Our BBB-penetrant oral PROTAC molecules can be optimized to show degradation of the LRRK2 protein in the brain across species (see Keystone Presentation) and broadly biodistributed to deep anatomic regions of the primate brain. As shown in the figure below, following oral administration the LRRK2 PROTAC -induced degradation of LRRK2 across the primate brain including the cortex, striatum, and cerebellum. These brain regions are important in neurodegenerative diseases and are difficult for biologic and genomic modalities to get to by intravenous or intrathecal dosing routes. Thus, further differentiating our BBB-penetrant PROTAC technology as a modality that can be orally administered and can uniformly biodistribute across the brain.
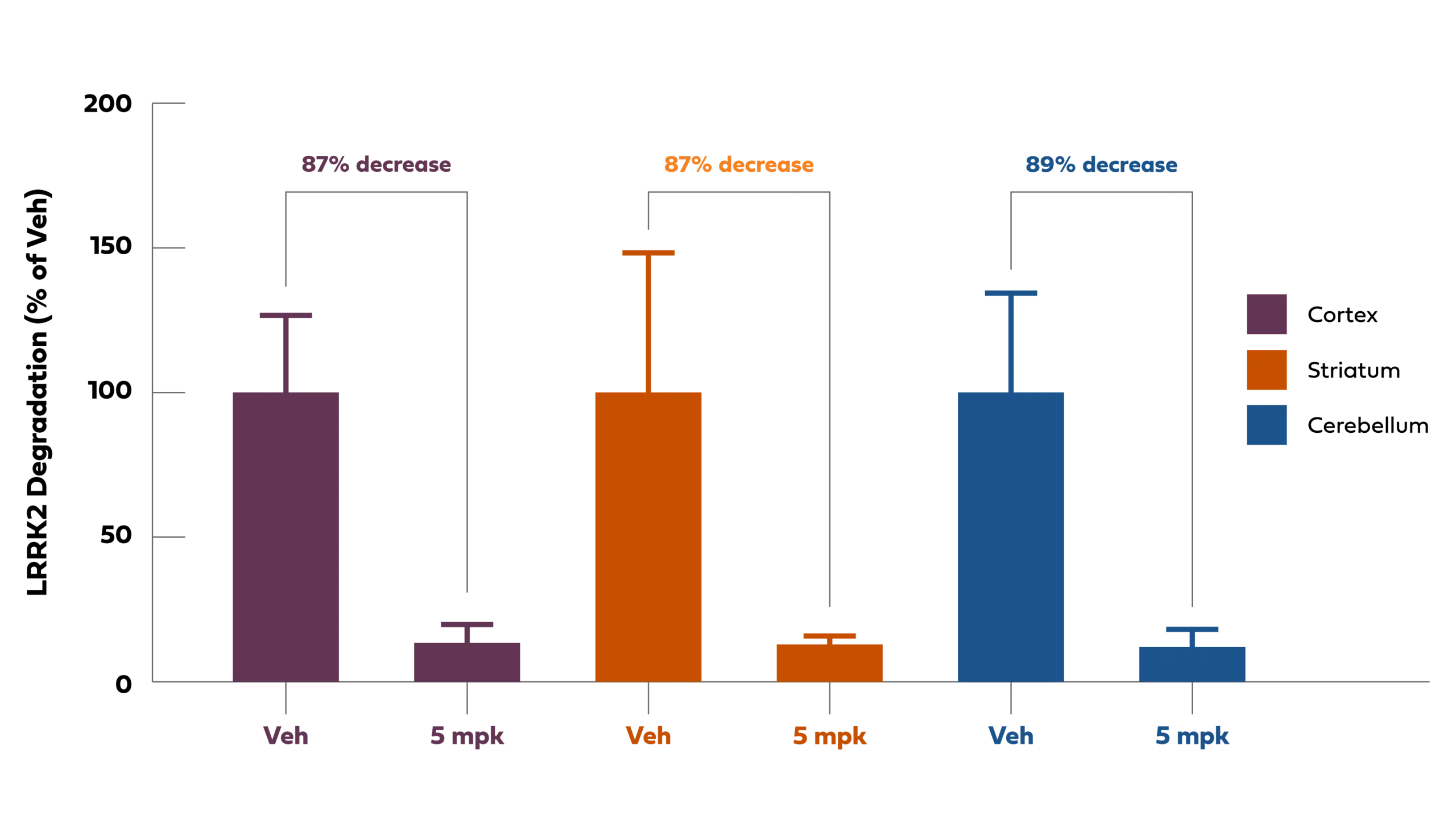
Chart Description: Oral PROTAC-induced LRRK2 degradation with biodistribution to deep brain regions
Preclinical Studies
In preclinical studies to date, Arvinas has demonstrated the ability to create PROTACs that can be dosed either orally or parenterally and cross the blood-brain barrier. Arvinas believes our ongoing research on PROTACs represent a promising new approach to treating diseases linked to pathogenic tau, alpha-synuclein, and other important targets previously considered “undruggable”.
Arvinas is progressing PROTAC programs to target three named protein targets (mutant huntingtin (mHTT), tau, and alpha-synuclein), and a number of undisclosed targets.
Huntington’s disease is caused by a mutation in the huntingtin (HTT) gene. PROTAC protein degradation has the potential to enable selective targeting of mutant HTT (mHTT) protein. Arvinas has identified ligands that, when incorporated into a PROTAC protein degrader molecule, induce potent degradation of mHTT. Protein degradation requires binding to both the target and E3 ligase. Our PROTAC molecule induces potent degradation of mHTT while sparing the protein produced by the wild-type HTT allele.
The tau protein is normally found in axons of healthy neurons of the central nervous system, and when it is mutated or under pathologic conditions, it has been implicated in a group of diseases known as tauopathies. Of these disorders, Alzheimer’s is an example. In this group of diseases, the pathologic tau protein dissociates from axons, negatively impacts neuronal function, becomes phosphorylated, acquires additional pathologic post-translational modifications, and forms insoluble clusters of aggregated proteins in the brain known as tangles. In preclinical studies, Arvinas has demonstrated that tau-targeting PROTACs, when dosed peripherally, degrade several different forms of tau, including phosphorylated forms, resulting in reduced insoluble aggregated tau in a mouse tauopathy model. The data shown below shows the relationship of PROTAC exposure in brain to reduction of PHF1 insoluble tau (aggregated tau is phosphorylated in a mouse tauopathy model).
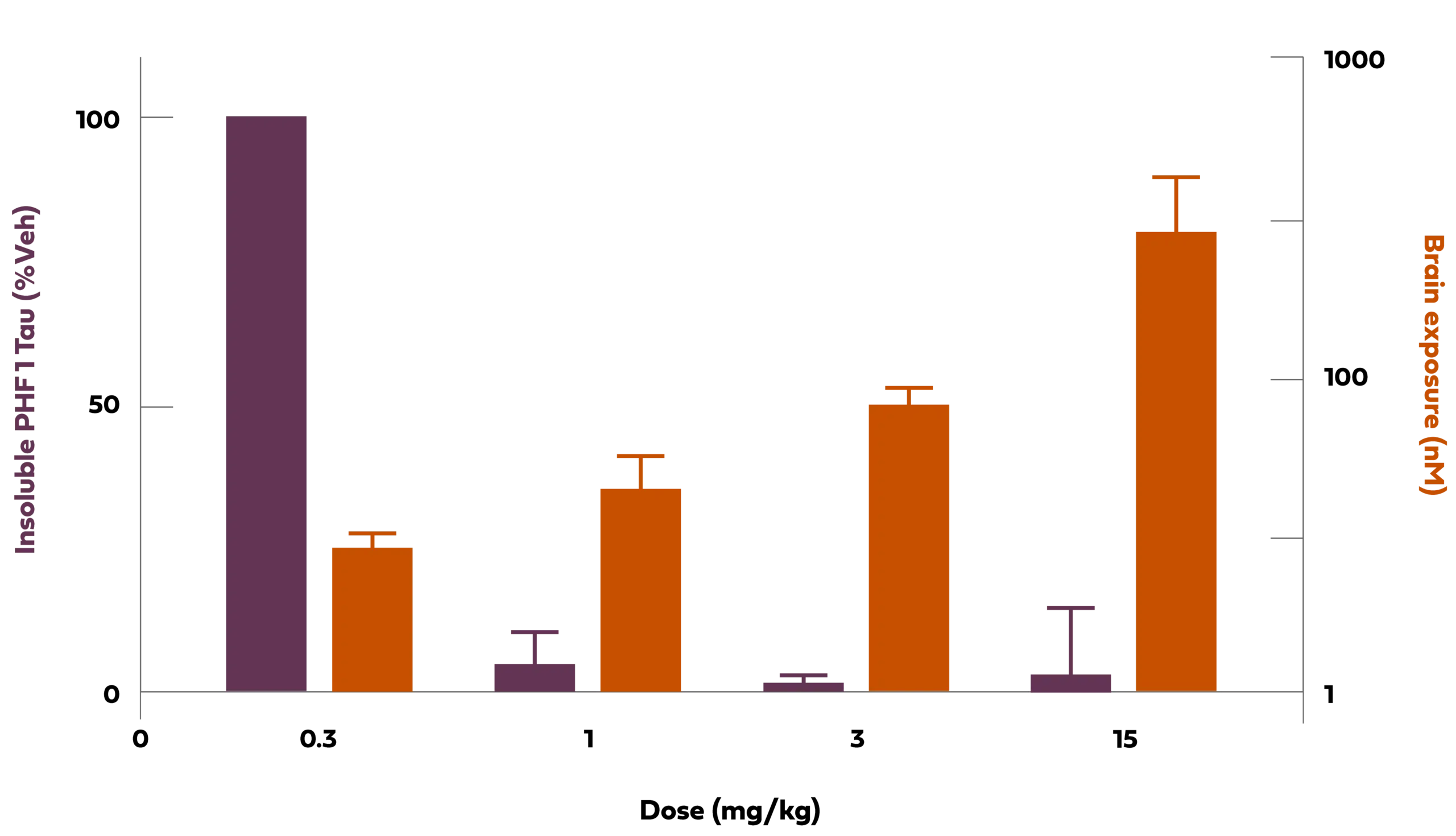
Chart Description: 1 Tg2508 is a murine pathologic tau model (P301L). nM: nanomolar;mg/kg:Milligrams per kilogram;veh: vehicle; PHF1 is an antibody direct against phosphorylated Ser396 and Ser404.
Alpha-synuclein (α-syn) is an abundant neuronal protein that localizes predominantly to presynaptic terminals within the brain. While the precise normal function of this protein is not fully understood, the pathologic aggregation of α-syn is implicated in synucleinopathies. The most common of these diseases is Parkinson’s.
Arvinas is working on approaches to degrade α-syn and remove this protein from neurons that lead to pathology in diseases such as Parkinson’s Disease, multiple system atrophy, and Lewy body dementia.
The agents listed above are currently under investigation. Their safety and effectiveness have not yet been established.
View our latest neuroscience-focused scientific presentation:
- Cacace, A. (2019, October 24). Discovery of Brain Penetrant PROTAC Degrader Molecules That Target Pathologic Tau and alpha-Synuclein Protein Species [1-22]. Presented at: Targeted Protein Degradation Summit. Boston, MA. https://s3.us-east-1.amazonaws.com/arvinas-assets.investeddigital.com/scientific-publications/Cacace_TPD-10-24-2019_final.pdf
- Pardridge, W. M. (2020). Blood-brain barrier and delivery of protein and gene therapeutics to brain. Frontiers in Aging Neuroscience, 11. https://doi.org/10.3389/fnagi.2019.00373
- Mazur, C., Powers, B., Zasadny, K., Sullivan, J. M., Dimant, H., Kamme, F., Hesterman, J., Matson, J., Oestergaard, M., Seaman, M., Holt, R. W., Qutaish, M., Polyak, I., Coelho, R., Gottumukkala, V., Gaut, C. M., Berridge, M., Albargothy, N. J., Kelly, L., Carare, R. O., … Verma, A. (2019). Brain pharmacology of intrathecal antisense oligonucleotides revealed through multimodal imaging. JCI insight, 4(20), e129240. https://doi.org/10.1172/jci.insight.129240
- Kouhi, A., Pachipulusu, V., Kapenstein, T., Hu, P., Epstein, A. L., & Khawli, L. A. (2021). Brain Disposition of Antibody-Based Therapeutics: Dogma, Approaches and Perspectives. International journal of molecular sciences, 22(12), 6442. https://doi.org/10.3390/ijms22126442
- Silva M. C., Ferguson F. M., Cai Q., Donovan K. A., Nandi G., Patnaik D., et al. (2019). Targeted degradation of aberrant tau in frontotemporal dementia patient-derived neuronal cell models. Elife 8, e45457. 10.7554/eLife.45457. https://pubmed.ncbi.nlm.nih.gov/30907729/
- Silva, M. C., Nandi, G., Donovan, K. A., Cai, Q., Berry, B. C., Nowak, R. P., Fischer, E. S., Gray, N. S., Ferguson, F. M., & Haggarty, S. J. (2022). Discovery and optimization of tau targeted protein Degraders enabled by patient induced pluripotent stem cells-derived neuronal models of Tauopathy. Frontiers in Cellular Neuroscience, 16. https://doi.org/10.3389/fncel.2022.801179
- Bond M.J. and Crews, C.M. (2021). PROTACs come of age: entering the third decade of targeted protein degradation. RSC Chem. Biol. https://pubs.rsc.org/en/content/articlepdf/2021/cb/d1cb00011j
- Bekes M., Langley, D. R., and Crews, C.M. (2022). PROTAC targeted Protein Degraders: the past is prologue. Nat. Rev. Drug Disc. https://www.nature.com/articles/s41573-021-00371-6
- Cacace, A. (2021, July 14). PROTAC Discovery Engine: Harnessing power of oral blood brain barrier penetrant degraders and new E3 ligases (1-22). Presented at Protein Society Meeting. Virtual. https://s3.us-east-1.amazonaws.com/arvinas-assets.investeddigital.com/scientific-publications/Angela-Cacace-Protein-Society-July-2021_2021-08-06-014908_ebim.pdf
- Cacace, A.M., Meredith, J., Bryce, D., Sparks, S., Kimmel., L., Kelly, K., Gregory, J., Matchett, M., Nickischer, D., Hendricson, A., Naumann, G., Kyne, R., Wilson., J., Corradi, J., Soto, L., Jeong, Y., Pizzano, J., Cadelina, G., Revell, D., Snyder, L., Berlin, M. (2022, November 12-16). Presented at Society for Neuroscience. San Diego, CA. Clearance of Pathologic Proteins in Neurodegeneration by oral PROTAC® Degrader Molecules.
- Burslem, G. M., Smith, B. E., Lai, A. C., Jaime-Figueroa, S., McQuaid, D. C., Bondeson, D. P., Toure, M., Dong, H., Qian, Y., Wang, J., Crew, A. P., Hines, J., & Crews, C. M. (2018). The Advantages of Targeted Protein Degradation Over Inhibition: An RTK Case Study. Cell chemical biology, 25(1), 67–77.e3. https://doi.org/10.1016/j.chembiol.2017.09.009 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5831399/#__ffn_sectitle
- Wang, X., Negrou, E., Maloney, M. T., Bondar, V. V., Andrews, S. V., Montalban, M., Llapashtica, C., Maciuca, R., Nguyen, H., Solanoy, H., Arguello, A., Przybyla, L., Moerke, N. J., Huntwork-Rodriguez, S., & Henry, A. G. (2021). Understanding LRRK2 kinase activity in preclinical models and human subjects through quantitative analysis of LRRK2 and pT73 Rab10. Scientific reports, 11(1), 12900. https://doi.org/10.1038/s41598-021-91943-4
- Zhao, Y., & Dzamko, N. (2019). Recent Developments in LRRK2-Targeted Therapy for Parkinson’s Disease. Drugs, 79(10), 1037–1051. https://doi.org/10.1007/s40265-019-01139-4
- Henderson, M. X., Sengupta, M., McGeary, I., Zhang, B., Olufemi, M. F., Brown, H., Trojanowski, J. Q., & Lee, V. M. Y. (2019). LRRK2 inhibition does not impart protection from α-synuclein pathology and neuron death in non-transgenic mice. Acta neuropathologica communications, 7(1), 28. https://doi.org/10.1186/s40478-019-0679-5
- Berwick, D.C., Heaton, G.R., Azeggagh, S. et al. (2019). LRRK2 Biology from structure to dysfunction: research progresses, but the themes remain the same. Mol Neurodegeneration 14, 49 https://doi.org/10.1186/s13024-019-0344-2
- Cacace, A. (2019, October 24). Discovery of Brain Penetrant PROTAC Degrader Molecules That Target Pathologic Tau and alpha-Synuclein Protein Species [1-22]. Presented at: Targeted Protein Degradation Summit. Boston, MA. https://s3.us-east-1.amazonaws.com/arvinas-assets.investeddigital.com/scientific-publications/Cacace_TPD-10-24-2019_final.pdf
- Change, C.-W., Shao, S., Mucke, L. (2021). Tau: enabler of diverse brain disorders and target of rapidly evolving therapeutic strategies. Science. 371(6532). https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8118650/pdf/nihms-1682624.pdf
- Rideout, H., Greggio, E., Kortholt, A., Nichols, R. J. (2022). Editorial: LRRK2—Fifteen Years From Cloning to the Clinic. Frontiers in Neuroscience. https://www.frontiersin.org/articles/10.3389/fnins.2022.880914
- Rocha, E.M., Keeney, M.T., Di Maio, R., De Miranda, B.R., Greenamyre, J.T. (2022). LRRK2 and idiopathic Parkinson’s disease. Trends in Neuroscience. 45(3), P224-236. https://www.cell.com/action/showPdf?pii=S0166-2236%2821%2900250-2
- Langston, R.G., Beilina, A., Reed, X., Kaganovich, A., Singleton, A.B., Blauwendraat, C., Gibbs, J.R., Cookson, M.R. (2022). Association of a common genetic variant with Parkinson’s disease is mediated by microglia. Science Translational Medicine. 14 (655). https://www.science.org/doi/10.1126/scitranslmed.abp8869
- Tabrizi SJ, Estevez-Fraga C, van Roon-Mom WMC, Flower MD, Scahill RI, Wild EJ, Muñoz-Sanjuan I, Sampaio C, Rosser AE, Leavitt BR. (2022) Potential disease-modifying therapies for Huntington’s disease: lessons learned and future opportunities. Lancet Neurol. (7):645-658. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7613206/pdf/EMS146360.pdf.
- Goedert M, Jakes R, Spillantini MG. The Synucleinopathies: Twenty Years On. (2017) J Parkinsons Dis. 7(s1):S51-S69. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5345650/pdf/jpd-7-jpd179005.pdf